Introduction
Species of the genus Vespa Linnaeus, 1758 (Hymenoptera: Vespidae) are apex predators of various arthropods and play critical regulatory roles in insect ecosystems (Brock et al., 2021; Richter, 2000). However, their potent venom and aggressive behavior pose risks to human safety (Choi et al., 2019), and certain species target honeybees, causing significant economic damage to apiculture (Norderud et al., 2021; Requier et al., 2023).
In recent years, the spread of Vespa species into regions such as Northeast Asia, Europe, and North America has raised concerns regarding their ecological, economic, and public health impacts (Lioy et al., 2022; Otis et al., 2023). The invasive alien species Vespa velutina Lepeletier, 1836, originally native to China, has expanded across Asia and Europe (Lioy et al., 2022), and was reported in the United States in 2023 (Hoebeke et al., 2024). Similarly, Vespa mandarinia Smith, 1852 drew international attention after its spread from Korea and Japan to North America in 2019 (Alaniz et al., 2021).
In invaded regions, various control efforts have been implemented to suppress hornet populations, reduce apiary damage, and mitigate broader ecological impacts (Beggs et al., 2011; Turchi & Derijard, 2018). However, most field-based control measures rely on direct collection using nets or hornet attractant traps, which are labor-intensive and offer short-term effectiveness (Beggs et al., 2011; Hsu et al., 2021; Lioy et al., 2020). Developing more sustainable and effective strategies requires a deeper understanding of Vespa biology, colony development, and foraging behavior; however, such ecological insights remain scarce for many species (Matsuura, 1984; Monceau et al., 2014).
Although Vespa species are frequently observed near apiaries (Chang et al., 1993; Choi & Kwon, 2015; Kim et al., 2024; Sim et al., 2014), only a limited subset directly threaten honeybees. This variation reflects interspecific differences in prey preferences and hunting behaviors (Matsuura, 1984; Matsuura & Yamane, 1990). In addition to predation, hornets may be drawn to sugar-rich resources, such as honey, pollen, and fermenting fruit (Couto et al., 2014; Paschapur et al., 2022), and abandoned hives or nearby areas serve as favorable nesting sites (Matsuura, 1971).
The seasonal activity of hornets is influenced by geographic and climatic variability, including local microclimates and annual weather fluctuations, which affect colony survival and growth (Choi & Kwon, 2015; Kim et al., 2020; Matsuura, 1991; Singha et al., 2023). The rise of invasive species has also led to shifts in the distribution of native hornets, introducing interspecific competition, and potentially altering colony cycles (Barbet-Massin et al., 2013; Choi et al., 2012a).
Early stage hornet colonies are often difficult to detect because of their high failure rates and small colony sizes (Spradbery, 1973), which limit the feasibility of direct observation. As an alternative, analyzing the seasonal occurrence of individuals collected using hornet attractant traps and distinguishing between overwintered queens and workers can provide valuable insights into colony development.
In the present study, a control method using insecticides was applied at the study apiary during fall (Hong & Jung, 2024; Kim et al., 2021), after which the abundance of certain Vespa species declined sharply. Although observational, these patterns highlight the potential influence of chemical control on hornet population dynamics and underscore the need for more rigorous experimental validation.
Here, we monitored Vespa activity over two years at an apiary in Daegu, South Korea, using hornet attractant traps. We examined the timing, duration, and frequency of the emergence of overwintered queens and workers across multiple species. These findings offer comparative insights into species-specific colony development and provide baseline ecological data for the development of more effective and ecologically informed hornet management strategies in apicultural environments.
Materials and Methods
Study site and trap configuration
This study was conducted at an apiary located near Palgongsan National Park in Daegu, South Korea (35°58’25.8”N, 128°39’48.2”E). The study area covered approximately 30 m². Five hornet attractant traps were deployed at 5-10 m intervals within the site (Kim et al., 2019). The attractant used in the traps was a mixture of plum extract and water at a 7:3 ratio. Given the relatively short distances between the traps and the wide foraging range of hornets, we assumed that the traps could potentially sample individuals from the same local population (Choi et al., 2012b). Therefore, the data from all traps were pooled for analysis.
Sampling schedule and frequency
For 2022, the survey focused on determining the emergence period of overwintered queens. Traps were installed on April 25 and collected seven times until July 30, when the queen activity ceased.
In 2024, to gain a more detailed understanding of species-specific queen emergence and seasonal dynamics of workers, traps were installed on April 7. Sampling was conducted 48 times between April 9 and October 22. During the expected emergence period of the new queens, traps were checked every 2-3 days to monitor the transition to the colony’s cooperative phase. After mid-July, when workers began to appear, the sampling intervals were extended to approximately two weeks (Table 1).
Species and caste identification
Among the insects collected from the traps, individuals belonging to the genus Vespa were isolated and their species and caste were identified. Identification was performed using the taxonomic key provided by Smith-Pardo et al. (2020).
Results
Species composition and abundance of Vespa queens and workers over two years
Over the survey period, 748 individuals representing eight Vespa species were collected from the apiary. Among these, V. velutina (N=256, 34.2%) and Vespa crabro Linnaeus, 1758 (N=217, 29.0%) were the most frequently collected species, followed by Vespa ducalis Smith, 1852 (N=131, 17.5%), V. mandarinia (N=70, 9.36%), and Vespa analis Fabricius, 1775 (N=40, 5.4%). Vespa dybowskii André, 1884, Vespa simillima Smith, 1868, and Vespa binghami du Buysson, 1905 were collected in substantially lower numbers, generally fewer than 10 individuals each, and were thus considered minor components of the hornet community at the site (Fig. 1).
In 2022, 186 overwintered queens representing six species were collected. V. velutina was the dominant species (N=102, 54.8%), followed by V. crabro (N=29, 15.6%) and V. mandarinia (N=20, 10.8%; Figs. 1, 2).
In 2024, 562 individuals from eight species will be collected. The most frequently collected species was V. crabro (N=188, 33.7%), followed by V. velutina (N=154, 27.6%) and V. ducalis (N=113, 20.3%). Among these, 171 overwintered queens from eight species were identified, with V. velutina being the most dominant (N=51, 29.8%), followed by V. crabro (N=43, 25.1%) and V. mandarinia (N=37, 21.6%; Fig. 2).
In 2024, 391 workers from seven species were collected. V. crabro accounted for the highest proportion (N=145, 37.1%), followed by V. velutina (N=103, 26.3%) and V. ducalis (N=102, 26.1%; Fig. 3).
Emergence timing of overwintered queens
Vespa velutina
In 2022, following trap installation on April 25, overwintered queens of V. velutina were predominantly collected during surveys on May 8 (N=51) and May 23 (N=38), indicating a sharp early peak in emergence. After May 30, the number of queens declined rapidly, with only 5-6 individuals collected intermittently in June. The last queen was collected on June 30.
In 2024, traps will be installed on April 7, and the first queen was collected on April 12. The highest number was recorded on April 29 (N=15), followed by consistent but declining collections of 3-4 individuals per survey until May 17, after which no further queens were collected. The final collection date for the queens was May 17.
These results indicate that V. velutina queens begin to emerge in early to mid-April, reach peak activity between late April and mid-May, and then decline sharply (Figs. 2, 4).
Vespa mandarinia
In 2022, the first queen was collected during the initial survey on May 8, and the queens continued to be collected in low numbers (≤6 individuals) until June 15, which marked the final collection date.
In 2024, the first queen was collected on April 9, with sporadic collections continuing until June 9. Considering that queen activity in 2022 may have already begun before the start of sampling, it is likely that V. mandarinia also began its emergence period from early to mid-April. In addition, three young queens were observed in the apiary on October 6, 2024 (Figs. 2, 4).
Vespa crabro
In 2022, the first V. crabro queen was collected on May 8, and a peak of 14 individuals was observed on May 23. Queens were collected at a steady rate (3-5 individuals per survey) until their final collection on July 2.
In 2024, the first queen was recorded on April 21 and was continuously collected in low numbers (2-3 individuals) until June 20. Although no individuals were collected for approximately one month thereafter, one additional queen was captured on July 18. These findings suggest that V. crabro begins to emerge in late April and remains active until late June, with some individuals persisting into mid-July (Figs. 2, 4).
Vespa analis
In 2022, the first V. analis queen was collected on May 8, and the final individual was recorded on May 30. In 2024, the first queen was collected on April 26, followed by intermittent captures (1-5 individuals) until June 12 without a clear emergence peak. One young queen was observed on September 22, 2024. These results suggest that V. analis typically begins to emerge in late April (Figs. 2, 4).
Vespa ducalis
Among the studied species, V. ducalis exhibited the most recent queen emergence. In 2022, the first queen was collected on May 30, with collection continuing until July 2 (N=18). In 2024, the queens were collected from June 12 to July 12 (N=9). Notably, on July 12, the queen was collected alongside a worker, suggesting an active colony formation. Several young queens were also observed from mid to late October, 2024. These findings indicate that V. ducalis exhibits delayed emergence and colony initiation compared to the other species (Figs. 2, 4).
Other species
V. dybowskii, V. simillima, and V. binghami were collected in low numbers with sporadic emergence patterns. V. binghami queens were collected on May 8 and 23, 2022, and on April 26 and May 19, 2024, indicating intermittent activity between late April and mid-May. V. dybowskii queens were collected on April 26, April 29, and June 9, 2024 (N=4 in total), suggesting occasional emergence between late and early June. V. simillima queens were collected only on April 29 and May 10, 2024 (N=5), indicating a limited and short emergence window between late April and early May.
Emergence timing of workers
Vespa velutina
In the 2024 survey, the first V. velutina worker was collected on July 10. No individuals were collected on the previous sampling date (July 8), indicating a sudden and synchronized onset of worker activity. Within two days, 30 individuals were collected, representing a sharp increase in the population. The number of workers temporarily declined over the following three weeks; however, another peak was recorded on August 7 with 25 individuals. From September to October, the number of individuals dropped sharply, with only four workers collected in each of the two months (Figs. 3, 4).
Vespa mandarinia
The first V. mandarinia worker was collected on July 18, 2024. Thirteen individuals were collected during the survey period. Overall, the abundance remained low, with no clear increase or peak (Figs. 3, 4).
Vespa crabro
Similar to V. velutina, V. crabro workers first appeared on July 10, 2024. The population gradually increased, peaking at 31 individuals on August 18. From late August to September, 10-20 individuals were consistently collected during each sampling period. However, by early October, worker occurrence dropped sharply, with the last collection occurring on October 6 (Figs. 3, 4).
Vespa analis
The first worker of V. analis was collected on August 7, 2024. However, fewer than five individuals were collected in August and September, and no clear pattern of increase was observed (Figs. 3, 4).
Vespa ducalis
Despite having a relatively late queen emergence, V. ducalis workers first appeared on July 12, 2024, around the same time as the other species. Four samples were collected on the same day. Fewer than ten individuals were collected in mid-August, after which the population began to increase. On September 22, 32 individuals were collected, which marked the peak in worker abundance. Worker numbers declined thereafter, with the last individuals (N=4) collected on October 6 (Figs. 3, 4).
Other species
For the remaining species, worker activity was minimal. V. dybowskii workers were collected sporadically between August and September. No V. simillima workers were collected during the survey period. V. binghami was represented by seven workers collected between early July and mid-September represented V. binghami. Overall, all three species exhibited limited sporadic worker activity.
Discussion
Vespa species observed in the apiary
Eight Vespa species were collected from the apiary during the study period. Among these, V. velutina was the most frequently collected species and was identified as the primary threat to apiaries. This species has now spread across South Korea and is the most prevalent hornet species near apiaries in most regions, contributing to widespread and significant damage (Choi & Kwon, 2015; Kim et al., 2025).
V. crabro and V. ducalis also appeared in relatively high numbers; however, these species tended to visit apiaries primarily to exploit floral resources rather than actively hunt honeybees. Consequently, their direct impact on apiculture is relatively minor (Matsuura & Yamane, 1990).
In contrast, although V. mandarinia was collected in smaller numbers, it poses a serious threat because of its tendency to engage in group attacks on beehives (Matsuura & Sakagami, 1973). Therefore, from a management perspective, V. velutina and V. mandarinia should be prioritized for control strategies in and around apiaries.
Emergence timing of overwintered queens
The sequences of overwintered queen emergence observed in the present study were V. mandarinia-V. velutina-V. crabro-V. analis-V. ducalis. V. mandarinia and V. velutina emerged the earliest in early April, followed by V. crabro and V. analis between late April and May. V. ducalis queens emerged in early June.
This emergence sequence closely aligns with previous observations in Japan by Matsuura (1984), who reported the queen emergence of V. mandarinia in April, V. analis and V. crabro in May, and Vespa tropica (Linnaeus, 1758), a species closely related to V. ducalis, between June 8 and 25.
Among the less frequently collected species, V. binghami, V. simillima, and V. dybowskii were primarily observed after late April. Notably, V. velutina exhibited an emergence pattern similar to that of V. mandarinia, with activity beginning in early April.
The overall advancement in emergence timing compared to earlier studies may be attributed to recent increases in average spring temperatures on the Korean Peninsula (An et al., 2011), which may accelerate the post-hibernation activity of overwintered queens (Spradbery, 1973).
Among all species, V. ducalis exhibited the latest colony initiation. This may reflect an adaptive strategy linked to its specialized ecological niche as a predator of paper wasps, with emergence timing synchronized with the development of its prey colonies (Archer, 1991; Matsuura, 1984; Starr, 1992).
From a methodological standpoint, the 2022 survey had wider intervals between site visits, limiting the precision of the emergence timing data. In contrast, the 2024 survey employed a shorter sampling interval, allowing for a more accurate tracking of emergence patterns.
Overall, most species ceased external activity from early to mid-June, after which queens were seldomly collected. This is likely due to the shift in colony dynamics around mid-June, when the first workers begin to emerge and the queens remain in the nest to focus on reproduction.
Seasonal emergence patterns of workers
This study examined the timing and abundance of hornet workers in an apiary. Most workers began to appear between early and mid-July, although in some species, the first individuals were not collected until mid-August. Worker abundance generally increased over time, peaking in August and September, followed by a sharp decline in October. This trend is consistent with previous reports on the life history of Vespa species, in which worker activity typically intensifies in mid-summer and lessens during autumn (Chang et al., 1993; Choi & Kwon, 2015; Matsuura & Yamane, 1990).
Interestingly, the current study showed a brief gap in worker collection between mid-June and early July. This may be attributed to the behavioral characteristics of early emerging workers, who are often smaller and less experienced in foraging. These individuals may avoid high-risk environments, such as apiaries, or escape from traps more easily because of their smaller size or less aggressive behavior (personal observation by Choi). Such traits could influence the frequency of their appearance at the apiary and the likelihood of their being collected by traps.
A particularly notable case was observed for V. velutina (Fig. 3). This species exhibited a sudden spike in worker abundance around July 10, with 30 individuals collected after a period of no capture. This sudden emergence coincided with approximately 250 mm of heavy rainfall recorded the day before and morning of sampling (KMA, 2024). It is likely that adverse weather conditions previously suppressed foraging activity, leading to delayed but concentrated post-rain emergence. Given the strong preference of V. velutina for honeybee predation (Norderud et al., 2021), such intensified activity near the apiary was expected.
However, these results were based on observations from a single apiary and may not reflect broader patterns of worker emergence across different landscapes. Factors such as topography, microclimate, vegetation, and local management practices also affect hornet activity. Thus, these findings should be interpreted with caution and should be supported by further studies across diverse geographical regions.
Implementation and effects of insecticide-based control
In this study, an insecticide-based control method targeting V. velutina was applied by the beekeeper at the apiary. The treatment was conducted beginning on July 25, using a method in which hornets were collected, treated with a neonicotinoid insecticide on the thorax and abdomen, and then released. The active ingredient used was clothianidin, a commercially available chloronicotinyl compound (International Organization for Standardization name: 3-[(2-chloro-1,3-thiazol-5-yl) methyl]-2-methyl-1-nitroguanidine; SG Hankooksamgong).
Immediately following the treatment, no sharp decline in trap collection was observed. According to previous studies, clothianidin, when applied at the recommended concentrations, can induce high mortality in adults within 6 hours and in larvae within 48 hours (Hong & Jung, 2024). Furthermore, this compound has been reported to spread throughout colonies within 24 hours, exerting a colony-wide effect (Kim et al., 2021). In theory, a noticeable population decline should have occurred within two days of application. However, as of August 7 sampling, 25 V. velutina workers were still being collected.
This delayed response may be due to the timing of the treatment, which coincides with the peak period of nest relocation in V. velutina (Diéguez-Antón et al., 2022). It is possible that newly arrived colonies that were not exposed to the insecticide subsequently established foraging routes for the apiaries. Hornet larvae are protected during pupation by cocoons, which form a sterile microenvironment that may prevent sufficient penetration by insecticides (Shabtai & Ishay, 1998). Given that pupation lasts approximately 16-19 days (Archer & Penney, 2012; Dong & Wang, 1989), and that clothianidin degrades naturally within approximately one week, it is plausible that newly emerged adults developed after the insecticide’s effective period had passed.
Despite these factors, the sustained application of the insecticide by the beekeeper likely contributed to the cumulative control effects over time. The number of V. velutina workers declined markedly from late August onward and did not recover until mid-October. These results suggested that the insecticide-based method had at least moderate efficacy in suppressing V. velutina activity.
Clothianidin, a neonicotinoid, is widely regarded by local beekeepers as an effective control agent for V. velutina (Hong & Jung, 2024). The findings of this study support the potential utility of such methods in mitigating hornet-related damage to apiaries. However, the repeated use of insecticides raises concerns about their unintended impacts on non-target organisms and residual contamination within apiaries. Therefore, future efforts should prioritize the development of more sustainable and ecologically responsible control strategies.
Conclusion
This study confirmed that V. velutina was the most damaging hornet species in the surveyed apiary in Daegu, with V. mandarinia posing a significant threat due to its capacity for group predation. In contrast, V. crabro and V. ducalis were frequently observed, but posed minimal direct risk to honeybee colonies.
The emergence patterns of overwintered queens varied by species, with most queens ceasing external activity by mid-June, coinciding with the transition to worker production. Workers began emerging in mid-July, peaked in August and September, and declined sharply in October. Notably, a substantial reduction in V. velutina was observed following insecticide treatment, indicating the effectiveness of the control method.
Nevertheless, owing to the potential risks of repeated insecticide use, particularly with respect to non-target species and environmental safety, future research should explore more sustainable and eco-friendly approaches to hornet management. This study provides baseline ecological data on hornet phenology and behavior in apiary environments that can inform the development of effective targeted management strategies.
Author Contributions
Conceptualization: Moon Bo Choi, Jaehee Kim. Data curation: Jaehee Kim. Formal analysis: Jaehee Kim. Funding acquisition: Moon Bo Choi. Investigation: Moon Bo Choi, Jaehee Kim. Methodology: Moon Bo Choi. Project administration: Ohseok Kwon, Moon Bo Choi. Resources: Moon Bo Choi. Software: Moon Bo Choi, Jaehee Kim. Supervision: Ohseok Kwon, Moon Bo Choi. Validation: Ohseok Kwon, Moon Bo Choi. Visualization: Moon Bo Choi, Jaehee Kim. Writing – original draft: Jaehee Kim. Writing – review & editing: Moon Bo Choi, Jaehee Kim.
References
Lioy, S., Bergamino, C., and Porporato, M. (2022). The invasive hornet Vespa velutina: distribution, impacts and management options. CABI Reviews, 2022. https://doi.org/10.1079/cabireviews202217030
Figures and Table
Fig. 1
Species and number of Vespa individuals collected through trapping over two years at the Daegu apiary.
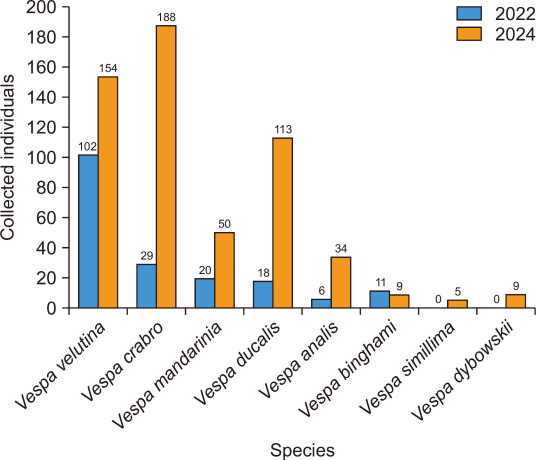
Fig. 2
Number of Vespa overwintered queens collected at the Daegu apiary from April to July in 2022 (yellow line) and 2024 (black line).
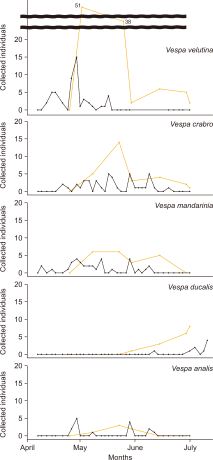
Fig. 3
Number of Vespa workers collected at the Daegu apiary from July to October in 2024 (the red X mark indicates the start of insecticide application on July 25, 2024).
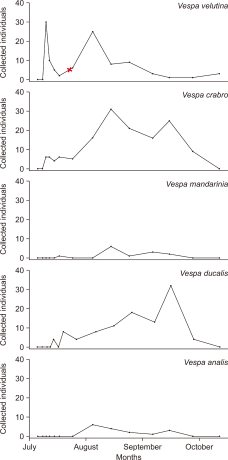
Fig. 4
Seasonal occurrence patterns of overwintered queens from five major Vespa species collected over two years at the Daegu apiary.
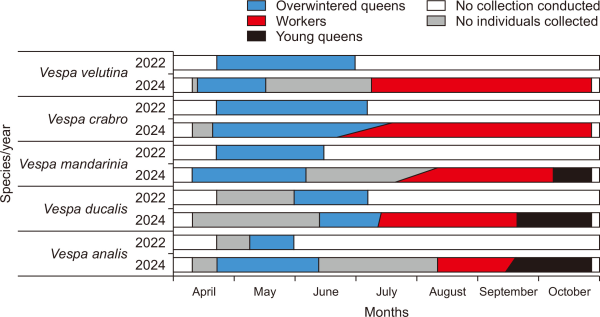
Table 1
Trap collection dates for Vespa sampling at the Daegu apiary from April to October in 2022 and 2024
| Month | Year | Day | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| April | 2022 | 25* | - | - | - | - | - | - | - | - | - | - | - | - |
| 2024 | 07* | 9 | 12 | 15 | 17 | 21 | 24 | 26 | 29 | - | - | - | - | |
| May | 2022 | 8 | 23 | 30 | - | - | - | - | - | - | - | - | - | - |
| 2024 | 1 | 3 | 6 | 8 | 10 | 13 | 15 | 17 | 19 | 22 | 24 | 27 | 29 | |
| June | 2022 | 15 | 30 | - | - | - | - | - | - | - | - | - | - | - |
| 2024 | 1 | 3 | 5 | 7 | 9 | 12 | 14 | 17 | 20 | 23 | 26 | 28 | - | |
| July | 2022 | 2 | 30 | - | - | - | - | - | - | - | - | - | - | - |
| 2024 | 2 | 5 | 8 | 10 | 12 | 15 | 18 | 26 | - | - | - | - | - | |
| August | 2024 | 7 | 18 | 29 | - | - | - | - | - | - | - | - | - | - |
| September | 2024 | 12 | 22 | - | - | - | - | - | - | - | - | - | - | - |
| October | 2024 | 6 | 22 | - | - | - | - | - | - | - | - | - | - | - |