Introduction
Anthropogenic greenhouse gas emissions have significantly altered the global climate. Global climate change is increasingly threatening human societies and natural ecosystems (Malhi et al., 2020). According to the 2018 Intergovernmental Panel on Climate Change (IPCC) Special Report on 1.5°C, an increase in the global temperature beyond 1.5°C would have severe climate change impacts such as drought, floods, heat waves, and rise in sea-level, which are detrimental to humankind and ecosystems (IPCC, 2018). In particular, sessile plants are the most closely exposed to climate change. Extreme weather events, combined with soil contaminants and various man-made pollutants, have greatly amplified the frequency and intensity of stressors that affect plants (Jing et al., 2024; Zandalinas et al., 2021). In addition, certain environmental conditions have been found to enhance the vulnerability of plants to attack by various pathogens or insects (Cohen & Leach, 2020; Desaint et al., 2021; Hamann et al., 2021). Therefore, it is necessary to conduct various studies, including research on resistance mechanisms, to conserve and restore plants vulnerable to climate change.
Climate change leads to the formation of vulnerable ecosystems through loss of biodiversity. Vulnerable ecosystems mainly focus on areas susceptible to climate change, such as islands, coasts, and subalpine zones (Hardy, 2003). Vulnerable subalpine ecosystems are located in high-altitude areas and highly sensitive to climate change. Plants in the subalpine regions are particularly threatened by limited opportunities for upslope migration (Bell et al., 2014; Koo et al., 2017; Randin et al., 2009). Since the late 1990s, global forest mortality and tree death rates have increased rapidly owing to high temperatures and droughts caused by climate change (Allen et al., 2010). Moreover, extensive tree mortality is often linked to prolonged drought stress, which affects coniferous forests in subalpine zones (Allen et al., 2010). Therefore, plants distributed in the subalpine zone are more affected by environmental stresses than those in low-mountain zones, and their habitats may decrease, or, in severe cases, become extinct. In addition, relict species in the subalpine zone are also significantly affected, with their distribution range likely to shrink because of the accelerating impact of climate change.
Korean fir (Abies koreana), a relict and endemic tree species, is distributed in the subalpine zone of mountain summits, such as Mts. Halla, Jiri, and Deogyu, in southern South Korea (Lee et al., 1982). It is a valuable tree species for ornamental purposes, and the essential oil extracted from it has showed antioxidant, anti-inflammatory, and antimicrobial activities (Jeong et al., 2007; Lee & Hong, 2009; Woo et al., 2008; Yoon et al., 2009). However, the population of A. koreana has been declining because of global warming, and A. koreana has been classified as a threatened species by the International Union for Conservation of Nature (IUCN) (Kim et al., 2011; Woo, 2009). Further, the regional decline in A. koreana is believed to be caused by the combined effects of global warming and other environmental stresses (Koo et al., 2001; Woo, 2009). In particular, the A. koreana population on Mt. Halla has a limited chance of receiving migrants from mainland populations (Koo et al., 2017). The combined effects of global warming and geographical isolation could hasten the decline in A. koreana forests on Mt. Halla. Therefore, studies on conservation and restoration are urgently needed to reduce the decline in A. koreana populations.
We previously performed a comprehensive transcriptomic analysis of A. koreana under various environmental stresses, such as elevated CO2, heat, and drought, to elucidate the transcriptomic alterations associated with these stresses (Hwang et al., 2018; 2019; Park & Hwang, 2024). These results have greatly deepened our understanding of how A. koreana responds to elevated CO2, heat, and drought stress at the molecular level, via various environmental response pathways. In addition, the genes isolated from the transcriptome RNA-seq dataset have been used to assess A. koreana’s vulnerability to various environmental stresses under natural conditions (Kim et al., 2020; Park et al., 2024). These results suggest that the genes used as diagnostic markers could effectively assess the ecological vulnerability of A. koreana to various environmental stresses and contribute to conservation efforts in subalpine ecosystems. In this study, we analyzed the cone fidelity of A. koreana districts containing ecologically vulnerable (Yeongsil [YS]) and stable regions (Nambyeok [NB] and Nambyeok-Baeknokdam [N-B]) on Mt. Halla. The cone fidelity of the ecologically vulnerable regions was also lower than that of the stable regions. These results imply that various efforts are required to conserve vulnerable subalpine ecosystems, including A. koreana, to ensure the survival and succession of future generations within these ecosystems.
Materials and Methods
Cone collections of Abies koreana by districts in Mt. Halla
Immature cones of A. Koreana were collected from each of the three sites of ecologically vulnerable (YS) and stable (NB and N-B) regions on Mt. Halla (Fig. 1). These sites are located 1,500 m above sea level on Mt. Halla. The YS region represents a relatively accessible yet vulnerable ecosystem, where the decline in A. koreana is observable, whereas the NB and N-B regions represent the most well-preserved ecological regions of Mt. Halla, offering substantial value for scientific research because of their pristine environmental conditions and biodiversity. The cones were transported to the laboratory and stored at –4°C before the samples were used.
Identification of embryos in immature cones of Abies koreana
For seed sterilization, the wings were removed and then rinsed with sterilized distilled water (DW). Seeds were sterilized in an ethanol solution (70% ethanol:DW=3:1) for 5 minutes and rinsed once with sterilized DW. Subsequently, the seeds were treated with 10% hydrogen peroxide solution for 5 minutes and then rinsed with sterilized DW. The seeds were then shaken for 30 minutes in a solution containing 5% sodium hypochlorite-based commercial bleach (Yuhan Clorox Co., Seoul, Korea) mixed with Triton X-100 (Bioshop, Burlington, ON, Canada) (bleach:DW:Triton X-100=49.95:49.95:0.1), and rinsed three times with sterilized DW. The seeds were then treated with 50 mg/L cefoxime (PhytoTech LABS, Lenexa, KS, USA) for 3 minutes to remove bacterial contamination, followed by five rinses with sterilized DW. Sterilized seeds were placed on a sterile Whatman filter paper (Cytiva, Wilmington, DE, USA), and immature embryos were extracted layer-by-layer from the outer seed coat and observed under an liquid crystal display (LCD) digital microscope (Gasworld, Seoul, Korea).
Statistical analysis of cone fidelity by districts of Abies koreana in Mt. Halla
Cones of A. koreana from each of the three ecologically vulnerable (YS) and stable (NB and N-B) regions on Mt. Halla were used and analyzed to determine cone fidelity in A. koreana embryos. All experiments were repeated at least three times for the quantitative analysis. Group comparisons were conducted using one-way analysis of variance with Excel's Data Analysis Toolpak (Microsoft, Redmond, WA, USA). When statistical significance was observed (P<0.05), post hoc analyses were performed. For comparisons between two groups, an F-test followed by a two-tailed independent t-test was conducted in Excel, and results with P<0.001 were denoted as ***.
Results and Discussion
Identification of the sampling sites in Yeongsil, Nambyeok, and Nambyeok-Baeknokdam regions of Mt. Halla
Designated as a natural reserve in 1966, Mt. Halla became Hallasan National Park in 1970, was recognized as a United Nations Educational, Scientific and Cultural Organization (UNESCO) Biosphere Reserve in 2002, and was listed as a World Natural Heritage Site in 2007. Mt. Halla is the highest mountain at 1,950 m in South Korea and has the widest distribution area of A. koreana. However, the declining patterns of A. koreana on Mt. Halla are the most serious and are ongoing because of climate change and various environmental factors caused by global warming. To compare the cone fidelity of the vulnerable and stable regions on Mt. Halla, cones of A. koreana were collected from the YS, NB, and N-B regions. The YS region is ecologically vulnerable, whereas the NB and N-B regions are relatively stable (Park et al., 2024). Therefore, three plant cones of A. koreana from each region of Mt. Halla were used to analyze cone fidelity (Fig. 1). Each region clearly showed the regional patterns of both vulnerable and stable sites (Fig. 2). In addition, geographic sites of the plants for cones are located in YS (latitude; 33.358495-33.358847 and longitude; 126.507552-126.508566), NB (latitude; 33.357779-33.357816 and longitude; 126.523535-126.523692), and N-B (latitude; 33.356223-33.356670 and longitude; 126.534911-126.535163) (Table 1). Thus, the cones of these plants were subjected to cone fidelity analysis in the ecologically vulnerable and stable regions of A. koreana on Mt. Halla.
Analysis of cone fidelity from Abies koreana in Mt. Halla
The cones of A. koreana on Mt. Halla on Jeju Island are well known for their distinctive and ornamental cones. It grows upright and stands vertically on branches, unlike many other conifers whose cones hang downward. The cones were relatively small, typically 5-7 cm in diameter), cylindrical, and tightly scaled. After maturation, the cones disintegrate and shed their seeds without falling to the ground in whole cone conditions, which is a characteristic trait of Abies species. However, cone fidelity is affected by various biotic and abiotic stressors caused by global warming. The observed damage rate appears to be sufficiently high to reduce the success of seed dispersal, which might ultimately impede subsequent seedling establishment and forest regeneration. To analyze the fidelity of the cones, immature cones were cut with a knife, and the embryos were extracted and separated using immature cones from ecologically vulnerable (YS) and stable (NB and N-B) regions on Mt. Halla. Thereafter, the embryos within the cone samples were examined using an LCD optical microscope and classified as either full or empty embryos (Fig. 3). Embryos of A. koreana were confirmed and cone fidelity was statistically calculated.
Comparison of cone fidelity by districts from Abies koreana embryos
We previously isolated candidate genes to assess ecological vulnerability using a comprehensive transcriptomic analysis of A. koreana under various environmental stressors, including elevated CO2, heat, and drought (Hwang et al., 2018; 2019; Park & Hwang, 2024). In addition, these genes have been confirmed for field applications to determine the ecological vulnerability of A. koreana under natural conditions (Kim et al., 2020; Park et al., 2024). These results strongly suggest that the YS region, located southwest of Mt. Halla, is an ecologically vulnerable region for the growth of A. koreana. In contrast, NB, which is situated south of the peak of Mt. Halla, is considered an ecologically stable region. Based on these results, we investigated the cone fidelity of A. koreana across different districts of Mt. Halla, including an ecologically vulnerable region (YS) and two relatively stable regions (NB and N-B) (Fig. 4). In each region, the cone fidelity of site 1 in the NB and N-B regions was approximately 2.1-2.4 times higher than that of the YS region (Fig. 4A). The cone fidelities in the NB and N-B regions at site 2 were approximately 4.1-4.7 times higher than those in the YS region (Fig. 4B). At site 3, the cone fidelity exhibited by the NB and N-B regions was approximately 4.7-5.1 times higher than that of the YS region. (Fig. 4C). Taken together, these results indicate that the cone fidelity of the ecologically vulnerable region (YS) was lower than that of the stable regions (NB and N-B), suggesting potential ecological disturbance or degradation in the YS region of Mt. Halla due to climate change.
In conclusion, global warming poses a critical threat to the survival of future generations, with particularly severe implications for ecologically vulnerable regions, such as islands, coasts, and subalpine regions. A. koreana, an endemic species found in the subalpine region of South Korea, is increasingly subjected to environmental stress caused by elevated winter temperatures and drought stress associated with climate change. Consequently, further research is required to understand species-specific adaptive responses to these environmental stresses under natural ecological conditions. In this study, we confirmed that the cone fidelity of the stable regions (NB and N-B) was approximately 2-5 times higher than that of the vulnerable region (YS) on Mt. Halla. These results imply that the cone fidelity of A. koreana is greatly affected by various environmental stresses on Mt. Halla, suggesting differential vulnerability to climate change. These findings provide a valuable foundation for developing conservation and restoration strategies for vulnerable subalpine ecosystems under ongoing climate change.
Author Contributions
Conceptualization: HCP. Data curation: HCP. Formal analysis: HCP, DYL. Funding acquisition: HCP. Methodology: HCP. Project administration: HCP. Supervision: HCP. Validation: HCP. Visualization: HCP, DYL. Writing – original draft: HCP. Writing – review & editing: HCP.
Funding
This work was supported by a grant from the National Institute of Ecology (NIE) funded by the Ministry of Environment (MOE) of the Republic of Korea (NIE-B-2025-15).
References
, , , , , , et al. (Eds.), Intergovernmental Panel on Climate Change (IPCC) (2018) Global Warming of 1.5°C: IPCC Special Report on Impacts of Global Warming of 1.5°C above Pre-industrial Levels in Context of Strengthening Response to Climate Change, Sustainable Development, and Efforts to Eradicate Poverty IPCC SPM: summary for policymakers, pp. 3-24
Figures and Table
Fig. 1
Photographs of sampling sites for Abies koreana cones in Mt. Halla. Images of (A) Yeongsil region, (B) Nambyeok region, and (C) Nambyeok-Baeknokdam region in Mt. Halla.
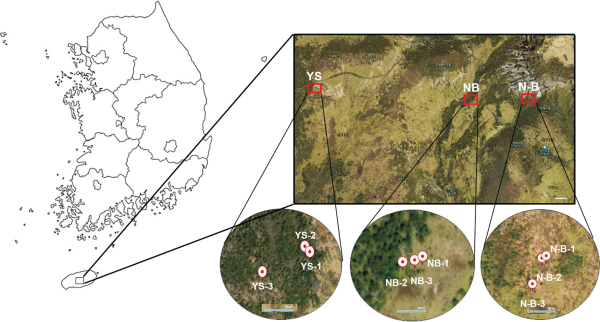
Fig. 2
Study areas for the sampling of Abies koreana cones in Mt. Halla. The regions marked with the red rectangular boxes represent YS, NB, and N-B regions of A. koreana in Mt. Halla, respectively. White scale bar indicates 100 m. YS, Yeongsil; NB, Nambyeok; N-B, Nambyeok-Baeknokdam.

Fig. 3
The process for analyzing the cone fidelity from Abies koreana embryos. Image of a healthy embryo of A. koreana (full). Image of an unhealthy embryo of A. koreana (empty).
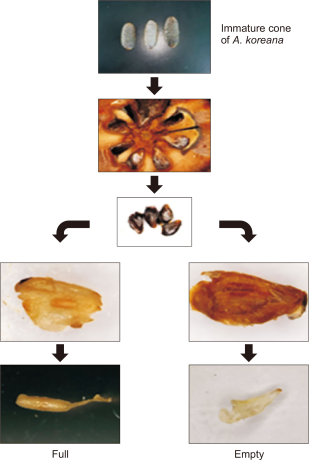
Fig. 4
Comparative analysis of cone fidelity by districts of Abies koreana in Mt. Halla. (A) Analysis of cone fidelity of A. koreana in YS, NB, and N-B regions at each site 1. (B) Analysis of cone fidelity in YS, NB, and N-B regions at each site 2. (C) Analysis of cone fidelity in YS, NB, and N-B regions at each site 3. Statistically significant differences relative to YS are indicated using asterisks (***P<0.001; Student’s t-test). YS, Yeongsil; NB, Nambyeok; N-B, Nambyeok-Baeknokdam.
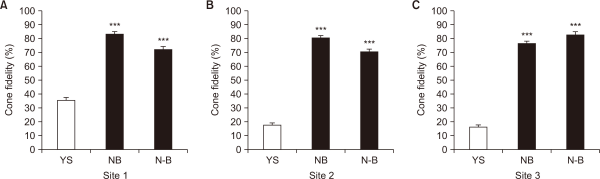
Table 1
Geographic information of sampling sites for Abies koreana cones from YS, NB, and N-B in Mt. Halla
| Site no. | Latitude | Longitude | Site no. | Latitude | Longitude | Site no. | Latitude | Longitude |
|---|---|---|---|---|---|---|---|---|
| YS-1 | 33.358772 | 126.508566 | NB-1 | 33.357816 | 126.523692 | N-B-1 | 33.356670 | 126.535163 |
| YS-2 | 33.358847 | 126.508462 | NB-2 | 33.357779 | 126.523535 | N-B-2 | 33.356662 | 126.535110 |
| YS-3 | 33.358495 | 126.507552 | NB-3 | 33.357792 | 126.523627 | N-B-3 | 33.356223 | 126.534911 |