Introduction
Coastal ecosystems are transitional zones where the land meets the sea, and where diverse environmental factors such as tidal cycles, salinity, moisture conditions, and soil characteristics intersect to form unique vegetation structures and functions (Bertness & Shumway, 1993; Lee & Ihm, 2004). Among these, salt marshes, reclaimed lands, freshwater wetlands, and sand dunes are subject to different intensities of disturbance and stress, and such differences provide key insights for understanding the distribution and ecological strategies of plant communities in these habitats (Álvarez-Rogel et al., 2007; Lee & Kim, 2018).
Grime’s CSR theory (Grime, 1974) explains plant survival strategies along three axes: Competitor (C), Stress-tolerator (S), and Ruderal (R). With the subsequent establishment of quantitative procedures by Hodgson et al. (1999) and Pierce et al. (2007; 2013), the CSR model has become widely applied as a methodological framework to interpret the functional responses of plant communities to environmental stress gradients (Negreiros et al., 2014).
In Korea, several studies have applied the CSR model to various coastal habitats. Son et al. (2020) analyzed soil factors and plant functional traits in salt marshes on the southwestern coast and showed that many halophytes adopted CR or SC strategies. Yang et al. (2021) confirmed the dominance of R and CR strategies in the early successional vegetation of the Saemangeum reclaimed land. Park et al. (2025) compared freshwater wetlands, sand dunes, salt marshes, and reclaimed lands in Muan, identifying distinct CSR strategies by habitat type and reporting that R/CR strategies were particularly prominent in freshwater wetlands. Choi (2022) performed canonical correspondence analysis (CCA) on tidal flats and salt marshes in the southwest coast, demonstrating close correlations between soil factors such as salinity, nitrogen, and organic carbon and plant strategies, and confirmed the prevalence of CR strategies in many species. Recently, Choi et al. (2024) further explored the habitat gradient–plant trait relationships, emphasizing the need for expanded studies across coastal–inland transitional zones.
However, most previous studies have focused on individual habitats, and few have provided integrated comparative analyses of multiple coastal habitats in Korea from the perspective of habitat connectivity (continuum). In other words, comprehensive understanding has not yet been fully established regarding how the stress adaptation of salt marshes, disturbance responses of reclaimed lands, rapid expansion of freshwater wetlands, and intermediate strategies of sand dunes are interconnected along a single environmental gradient.
Based on previous findings, the present study aims to analyze the species composition and functional traits of salt marshes, reclaimed lands, freshwater wetlands, and sand dunes along the southwestern coast of Korea in an integrated manner, and to compare the distribution of CSR strategies. Through this, we seek to identify the functional adaptations of plant communities along disturbance and stress gradients, and to provide foundational data for the conservation and restoration of coastal and inland transitional ecosystems.
Materials and Methods
This study analyzed and utilized the results of previous works by Choi et al. (2024), Lee et al. (2020), Park et al. (2025), Son et al. (2020), Yang et al. (2021).
Study sites
Field surveys were conducted in the southwestern coastal region of Jeollanam-do and the Saemangeum reclaimed area in Jeollabuk-do. The study sites comprised: (i) salt marshes subject to periodic tidal inundation (Son et al., 2020), (ii) reclaimed lands formed through artificial reclamation (Yang et al., 2021), (iii) sand dunes shaped by sand deposition and wind action (Lee et al., 2020), and (iv) freshwater wetlands characterized by freshwater-dominated environments (Park et al., 2025). These sites were selected to represent distinct gradients of salinity, water availability, disturbance intensity, and soil properties, providing a suitable basis for CSR strategy comparisons (Fig. 1, Table 1).
Plant functional traits
For the classification of CSR strategies, key functional traits were measured for dominant plant species in the study area (Hodgson et al., 1999; Pierce et al., 2013): leaf area (LA), leaf dry weight (LDW), leaf dry matter content (LDMC), specific leaf area (SLA), canopy height (CH), lateral spread (LS), flowering start (FS) and flowering period (FP).
From July to September between 2018 and 2025, plant samples were collected in the field, subsequently dried, pre-processed, and analyzed to determine C, S, and R scores for each species, using 1×1 m quadrats with three replicates per species.
Data analysis
CSR strategy proportions for each species were derived based on measured traits using the methods of Hodgson et al. (1999) and Pierce et al. (2013), and plotted on CSR ternary diagrams with Sigmaplot (Grafiti LLC, Palo Alto, CA, USA). The vegetation–environment relationship was assessed using CCA (Ter Braak, 1986; ter Braak & Šmilauer, 2018). Differences among habitats were tested with analysis of variance and multiple regression analysis (Table 2) (Hodgson et al. 1999; Son, 2019).
Results
Species distribution by habitat
Species composition differed clearly by habitat type. In salt marshes, halophytes such as Suaeda japonica, Salicornia europaea, Zoysia sinica, Phragmites communis, and Suaeda maritima were dominant, and community composition varied along the salinity gradient. In reclaimed lands, disturbance-adapted and naturalized species such as Chenopodium album var. centrorubrum, Echinochloa crus-galli, Setaria viridis, Sonchus brachyotus, Ambrosia artemisiifolia, Erigeron canadensis, and Trifolium repens were widely distributed. In sand dunes, dune-specialized species such as Calystegia soldanella, Carex kobomugi, and Ischaemum anthephoroides were found. Freshwater wetlands were dominated by aquatic and wetland plants such as Nelumbo nucifera, Ceratophyllum demersum, Oenanthe javanica, and Typha orientalis (Table 3).
Competitor, Stress-tolerator, and Ruderal strategy distribution and functional traits
Across all habitats, LA, LDW, LDMC, SLA, CH, LS, and FS/FP were measured to derive CSR strategies.
(i) Salt marshes: Plants generally exhibited low SLA and high LDMC, indicating strong resource-conserving traits. Species such as P. communis with high CH had competitive advantages for light capture. Overall, these communities showed CR or SC strategies. (ii) Reclaimed lands: Disturbance-adapted and naturalized species exhibited high SLA, low LDMC, and early, short flowering, allowing rapid adaptation to disturbance-prone environments. R/CR strategies predominated. (iii) Sand dunes: Plants showed well-developed LS and intermediate SLA and LDMC, adapting to dry, nutrient-poor, shifting sandy conditions. CR/CSR strategies were dominant. (iV) Freshwater wetlands: Large aquatic plants exhibited large LA, high CH, and high SLA, enabling rapid growth and light acquisition above the water surface. These communities were dominated by R/CR strategies (Fig. 2).
Integrated results
Along the habitat continuum, SC/CR strategies were dominant in salt marshes, R/CR strategies in reclaimed lands, CR/CSR strategies in sand dunes, and R/CR strategies in freshwater wetlands. The central axis of CSR shifted from stress-tolerant strategies (C/S) toward ruderal strategies (R), and then toward intermediate competitor–ruderal strategies (CR/CSR). This indicates that water conditions and disturbance intensity, mediated through functional traits and strategies, shape species composition and community structure.
Discussion
This study examined species composition and functional traits of plant communities across salt marshes, reclaimed land, sand dunes, and freshwater wetlands along the southwestern coast of Korea, applying Grime’s (1974) CSR theory to compare adaptive strategies among habitats. The findings revealed that distinct community structures and functional traits were closely associated with habitat-specific environmental drivers, including salinity, disturbance, water availability, and nutrient conditions.
In salt marshes, elevated salinity and tidal flooding promoted the dominance of halophytes such as S. japonica and S. europaea, which exhibited stress-tolerant or CR/SC strategies. Reclaimed land, shaped by unstable salinity and intense anthropogenic disturbance, was dominated by ruderal and naturalized species, primarily expressing R/CR strategies. Freshwater wetlands, under low-salinity, high-moisture, and nutrient-enriched conditions, supported large aquatic plants capable of rapid growth and dispersal, also reflecting R/CR strategies. In contrast, sand dunes, characterized by arid, oligotrophic conditions and sand mobility, favored dune specialists such as C. kobomugi and C. soldanella, which adopted intermediate CR/CSR strategies.
Overall, these results underscore the pivotal role of stress and disturbance gradients in shaping habitat-specific CSR strategies and functional traits. Furthermore, the coexistence of diverse adaptive strategies across the coastal–inland transition zone illustrates a critical ecological mechanism that sustains species diversity and functional stability in dynamic coastal ecosystems.
Conclusion
Overall, the findings highlight the pivotal role of stress and disturbance gradients in shaping habitat-specific CSR strategies and functional traits. The coexistence of diverse adaptive strategies across the coastal–inland transition zone illustrates a fundamental ecological mechanism that underpins species diversity and functional stability in dynamic coastal ecosystems.
This study thus provides a comprehensive comparison of habitats along the southwestern coast of Korea, revealing distinct species compositions and CSR strategies associated with specific environmental conditions. These results offer valuable insights into how transitional ecosystems sustain themselves through functional adaptation under accelerating climate change and anthropogenic disturbance.
In particular, CSR strategies exhibited a directional shift along the habitat continuum-from stress tolerance to disturbance adaptation, and further toward intermediate forms integrating competition, stress, and disturbance. This pattern underscores the importance of designing restoration and management strategies that are tailored to local environmental gradients and the functional traits of resident plant communities.
Future research should incorporate long-term monitoring and advanced quantitative approaches such as RLQ and fourth-corner analyses to refine our understanding of the linkages among environment, plant traits, and species composition. Such efforts will not only enhance ecological knowledge of the southwestern coast of Korea but also provide a robust scientific foundation for the conservation and restoration of transitional ecosystems worldwide.
Author Contributions
Data curation: DHS. Funding acquisition: JWK. Methodology: DHS. Supervision: DHS. Visualization: JC. Writing – original draft: DHS. Writing – review & editing: JC, JWK.
Funding
This Research was supported by Research Funds of Mokpo National University in 2024.
Figures and Tables
Fig. 1
Map of study areas. The red circles represent salt marshes, the yellow triangles represent reclaimed lands, the green squares represent sand dunes, and the orange pentagons represent freshwater wetlands.
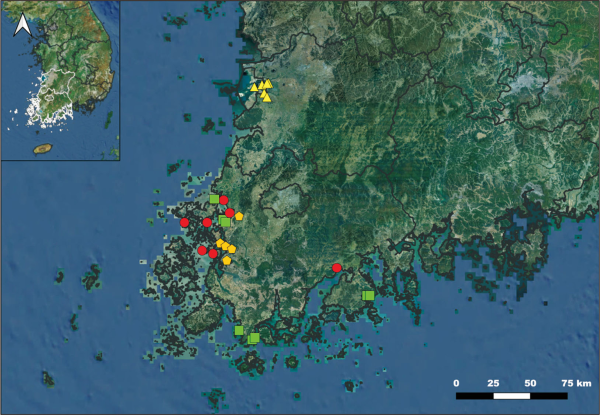
Fig. 2
CSR strategies of plant communities at each study site. (A) Salt marsh. SC/CR strategies were dominant. (B) Reclaimed lands. R/CR strategies prevailed. (C) Sand dune. CR/CSR strategies were dominant. (D) Freshwater wetland. R/CR strategies were predominant. C, Competitor; S, Stress-tolerator; R, Ruderal.
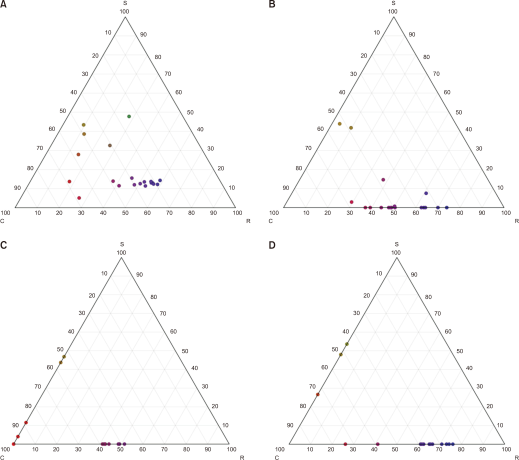
Table 1
Geographic coordinates of the study areas
| Type | Local name | Latitude | Longitude |
|---|---|---|---|
| Salt marshs | Haksan-ri | N 35.1592 | E 126.3783 |
| Haeun-ri | N 35.0718 | E 126.4523 | |
| Jeungdong-ri | N 34.9980 | E 126.1721 | |
| Songhyun-ri | N 34.9947 | E 126.3528 | |
| Sinjang-ri | N 34.8373 | E 126.3651 | |
| Daecheon-ri | N 34.8580 | E 126.2679 | |
| Haepyung-ri | N 34.7447 | E 127.2125 | |
| Sand dunes | Muan | N 35.0383 | E 126.4191 |
| Pyeongsan-ri | N 35.0276 | E 126.4237 | |
| Songseok-ri | N 35.1500 | E 126.3352 | |
| Songji | N 34.3656 | E 126.5069 | |
| Namyeol I | N 34.5792 | E 127.4767 | |
| Namyeol II | N 34.5796 | E 127.4864 | |
| Buldeung | N 34.3221 | E 126.5598 | |
| Sagumi | N 34.3204 | E 126.5809 | |
| Reclaimed lands | Seonyeon-ri | N 35.8904 | E 126.6362 |
| Wolyeon-ri | N 35.8943 | E 126.7367 | |
| Jeungseok-ri | N 35.9069 | E 126.7947 | |
| Hakdang-ri | N 35.9016 | E 126.7816 | |
| Eunpa-ri | N 35.8090 | E 126.7627 | |
| Yeonpo-ri | N 35.7554 | E 126.7826 | |
| Freshwater wetlands | Ogam riverside ecological park | N 34.8023 | E 126.4461 |
| Hoesan white lotus pond | N 34.8624 | E 126.5245 | |
| Yonggyecheon | N 34.9087 | E 126.4242 | |
| Hampyungcheon | N 34.0414 | E 126.5266 | |
| Gamdon reservoir | N 34.8815 | E 126.4705 |
Table 2
A definitions of the predictor variables used the CSR allocation procedure
| Variable | Definition | ||
|---|---|---|---|
| Canopy height | Six-point classification | 1 | 1-49 mm |
| 2 | 50-99 mm | ||
| 3 | 100-299 mm | ||
| 4 | 300-599 mm | ||
| 5 | 600-999 mm | ||
| 6 | >999 mm | ||
| Dry matter content | Mean of percent dry matter content in the largest, fully hydrated, fully expanded leaves (%) | ||
| Flowering period | Normal duration of flowering period (mo) | ||
| Flowering start | Six-point classification | 1 | First flowering in March or earlier |
| 2 | in April | ||
| 3 | in May | ||
| 4 | in June | ||
| 5 | in July | ||
| 6 | in August or later, or before leaves in spring | ||
| Lateral spread | Six-point classification | 1 | 1: Plant short-lived |
| 2 | Loose tufted ramets radiating about a single axis, no thickened rootstock (in graminoids) | ||
| 2 | Compactly tufted about a single axis, no thickened rootstock (in non-graminoids) | ||
| 3 | Compactly tufted ramets appressed to each other at base (in graminoids) | ||
| 3 | Compactly tufted about a single axis, thickened rootstock present (in non-graminoids) | ||
| 4 | Shortly creeping, <40 mm between ramets | ||
| 5 | Creeping, 40-79 mm between ramets | ||
| 6 | Widely creeping, >79 mm between ramets | ||
| Leaf dry weight | Natural logarithm of mean dry weight in the largest, fully hydrated, fully expanded leaves (mg), plus 3 | ||
| Specific leaf area | Mean of area/dry weight quotient in the largest, hydrated, fully expanded leaves (mm²/mg) | ||
Table 3
Major plant species by habitat type on the southwestern coast of Korea, with ecological strategies and cover percentages
| Habitat type | Major species | Ecological characteristics/strategy | Dominance/cover (%) |
|---|---|---|---|
| Salt marsh | Zoysia sinica, Phragmites communis | Typical halophytes adapted to high salinity and tidal flooding. Low SLA, high LDM→SC/CR strategies | Z. sinica 100, P. communis 60 |
| Reclaimed land | Chenopodium album, Echinochloa crus-galli | Disturbance-adapted and naturalized species. High SLA, low LDMC, short life cycle→R/CR strategies | C. album 50, E. crus-galli 40 |
| Sand dune | Carex kobomugi, Calystegia soldanella | Adapted to dry, nutrient-poor, and shifting sand conditions. Well-developed root systems and lateral spread→CR/CSR strategies | C. kobomugi 50, C. soldanella 60 |
| Freshwater wetland | Nelumbo nucifera, Typha orientalis | Low salinity, high water content, nutrient-rich conditions. Large leaves and tall stature→R/CR strategies | N. nucifera 70, T. orientalis 50 |