Introduction
Recently, climate change has driven the decline of coniferous forests in high-altitude and subalpine regions worldwide, and over the past 20 years, the area of subalpine coniferous forests in Korea has decreased by approximately 25% (Cao et al., 2021; Kim et al., 2019; Woo, 2009). In Hallasan, monitoring since 2003 has revealed a severe decline, with average mortality rate exceeding 36.4% in 2019 (Jeong et al., 2023; Kim et al., 2017). If temperatures continue to rise, the distribution of coniferous forests in Korea is predicted to decrease and face the risk of extinction in the future (Adhikari et al., 2018; Lee et al., 2025). Coniferous species in forests are more sensitive to climate change than deciduous species (Cao et al., 2021). Specifically, the combination of high temperatures and drought during winter and early spring causes water deficits, a major cause of the decline and mortality of evergreen conifers such as the Korean fir (Abies koreana E. H. Wilson) (Chandrasekaran et al., 2025). Efforts to conserve and restore vulnerable species, such as the Korean fir, are urgently required (Adhikari et al., 2018; Chandrasekaran et al., 2025).
The Korean fir is an endemic conifer species of the Korean Peninsula and serves as a key indicator of climate change because of its vulnerability to rising temperatures and drought (Hong et al., 2025; Kim et al., 2020; Koo & Kim, 2020; Park et al., 2015). This species is experiencing severe population decline, habitat loss, and increased mortality, primarily due to climate change-induced drought stress and water imbalance (Kim et al., 2024; Lim et al., 2025; Park et al., 2024). A major obstacle to conservation efforts is the extremely low natural germination rate of seeds, which limits the effectiveness of traditional propagation methods (Hong et al., 2025; Park & Lee, 2025). Therefore, in vitro propagation techniques, such as somatic embryogenesis, have emerged as critical biotechnological tools for the mass production and genetic preservation of this endangered species (Jouini et al., 2025).
The success of plant tissue culture depends on several factors, including the origin of the explant, its physiological state, and the specific composition of the culture medium (Guo & Jeong, 2021). Research on the genus Abies has established a foundation for understanding the effects of medium nutrient composition and hormonal balance on callus induction (Nawrot-Chorabik, 2008). However, within a single species, physiological state can vary owing to genetic diversity or collection environment, directly affecting culture outcomes. Research on the combined influence of physiological differences based on intra-cone embryo position and site-specific environmental factors on callus induction is still lacking.
This study aimed to compare and elucidate the effects of the collection site within Hallasan Mountain and the position of seeds within the cone on the callus induction rate of immature Korean fir embryos. Our goal was to develop an effective in vitro propagation protocol for Korean fir and provide a scientific basis for establishing a conservation strategy for this endangered species.
Materials and Methods
Plant materials
Immature cones of Korean fir (A. koreana E.H. Wilson) were collected from Hallasan Mountain in July (Fig. 1). To ensure the use of healthy materials, cones with straight shapes and no visible signs of pest or disease were selected. Collection was conducted at four representative sites: Witse Oreum, Nambyeok, Nambyeok-Baengnokdam, and Baengnokdam. The geographical coordinates of the sites are listed in Table 1. All sites were located within the subalpine zone of Hallasan, at elevations ranging from approximately 1,450 to 1,700 m above sea level.
The Witse Oreum site represents an ecologically vulnerable area where a population decline in Korean fir has been observed, reflecting environmental stress. In contrast, the Nambyeok and Nambyeok-Baengnokdam sites are relatively stable and well-preserved habitats with high biodiversity and pristine environmental conditions, serving as valuable reference areas for ecological studies. The Baengnokdam site, located near the summit crater, was included because it represents one of the highest natural habitats for the Korean fir.
After collection, cones were placed in sterile polyethylene bags, transported to the laboratory under chilled conditions, and stored at 4°C until further use for embryo isolation and culture. Additionally, cone samples from the Nambyeok-Baengnokdam and Baengnokdam sites were collected by help of the Habitat Conservation Team of the Center for the Restoration of Endangered Species, National Institute of Ecology.
Seed sterilization
To sterilize the seeds, the outer seed coat was removed, and the seeds were washed once with sterile distilled water (DW). They were sterilized in an ethanol solution (70% ethanol: DW=3:1) for 5 minutes and washed with sterile DW. This was followed by a 5-minute sterilization in 10% hydrogen peroxide solution and a final wash with sterile DW. Next, the seeds were agitated for 30 minutes in a solution containing commercial bleach (5% sodium hypochlorite; Yuhan Clorox, Seoul, Korea) and TritonX-10 (Bioshop, Burlington, ON, Canada) at a ratio of 49.95:49.95:0.1 (bleach:DW:TritonX-10). The remaining solution was removed by washing thrice with sterile water. Finally, to eliminate bacteria, seeds were treated with 50 mg/L cefoxime solution (PhytoTech Labs, Lenexa, KS, USA) for 3 minutes and washed five times with sterile water.
Immature embryo excision and plating
Immature embryos were excised on sterile filter paper (Cytiva, Wilmington, DE, USA) using an LCD digital microscope (Gasworld, Seoul, Korea). The inner seed coat was removed layer by layer to extract the immature embryos, which were then plated onto four different media types. Cultures were incubated for 4 weeks in a chamber at 24°C in the dark and subcultured on the same medium until the explants developed (callus formation).
Statistical analysis
A chi-square (χ2) test of independence was performed to examine the effects of medium composition and embryo position within the cone on callus formation rate. This test was used to determine whether a statistically significant relationship existed between the two categorical variables. All statistical analyses were conducted using R (version 4.5.1; R Foundation for Statistical Computing, Vienna, Austria), and statistical significance was determined at P<0.05.
Results
Explants plated by collection site and cone position
Korean fir cones were collected from four regions on Hallasan Mountain: Witse Oreum, Nambyeok, Nambyeok-Baengnokdam, and Baengnokdam (Fig. 2A, B). To obtain embryogenic calluses from Korean fir, seeds were first isolated from immature cones and sterilized (Fig. 2C, D). The outer and inner coats of the sterilized seeds were removed to extract the immature embryos, which was then plated onto the medium (Fig. 2E, F).
To compare callus formation based on cone position, explants were plated by dividing the cone into top, middle, and bottom sections (Fig. 3). The number of seeds plated per collection area was 582 from Witse Oreum, 140 from Nambyeok, 1,165 from Nambyeok-Baengnokdam, and 112 from Baengnokdam. When analyzed by cone position, the largest number of immature embryos was plated from the “middle” section of cones collected at the Nambyeok-Baengnokdam site. Conversely, the fewest immature embryos were plated from the “bottom” sections of cones collected at the Nambyeok site. Variation in the number of plated immature embryos was due to the careful selection of cones in good physiological condition. A total of 1,999 samples were collected for culture (Table 2).
Callus formation by region and cone position
Embryogenic calluses were successfully obtained from the immature embryos of Korean fir collected from Witse Oreum and Nambyeok-Baengnokdam (Fig. 4). As shown in Table 3, the Witse Oreum site yielded six out of 582 explants (1.0%), whereas the Nambyeok-Baengnokdam area showed a substantially higher formation rate of 20.1%, with 234 callus lines developing from 1,165 explants. The difference in callus formation rates between these two areas is likely attributable to the difference in the health of the immature embryos in the cones, suggesting that the cones from the Nambyeok-Baengnokdam area were under optimal conditions. No callus formation was observed from samples collected from the Nambyeok and Baengnokdam areas.
When considering the specific cone sections, the Witse Oreum samples showed a 0.8% formation rate in the top section (two out of 252 explants) and a 2.0% formation rate in the middle section (four out of 204 explants). No calluses developed in the bottom section. In the Nambyeok-Baengnokdam samples, the top section had a 24.0% formation rate (79 calluses from 329 explants), the middle section had a 20.8% formation rate (122 lines from 587 explants), and the bottom section had a 13.3% formation rate (33 calluses from 249 explants). The highest callus formation rate was observed in the top section of the cones from the Nambyeok-Baengnokdam area. This suggested that immature embryos from this specific location were in the healthiest condition.
Statistical analysis of callus formation
To statistically evaluate the effects of cone position on callus formation, a chi-square (χ2) test of independence was performed. The analysis confirmed a statistically significant difference in callus formation rate depending on cone position (χ2=11.68, df=2, P=0.0029). Immature embryos collected from the middle portion of the cone showed the highest formation rate, with 126 successfully formed calluses. This value is higher than the expected count of 107.45. In contrast, immature embryos from the bottom portion of the cone exhibited a notably lower rate, with only 33 calluses formed, compared to the expected count of 52.23 (Fig. 5). These results indicate that the location within the cone influences callus formation, with immature embryos situated at the bottom of the cone being less favorable for callus formation.
Discussion
In the present study, we successfully induced callus formation from immature embryos of Korean fir collected from four different sites on Hallasan Mountain. Our findings suggest that the location of the embryo within the cone and collection site significantly influences the callus formation rate.
The significant variation in callus formation rates among collection sites, with the Nambyeok-Baengnokdam area showing a substantially higher success rate than Witse Oreum and no callus formation from the Nambyeok and Baengnokdam sites, is likely attributed to the overall health and physiological conditions of the parent trees. This highlights the paramount importance of the health and genetic background of the source material for successful in vitro culture, and suggests that future studies should investigate these factors in greater detail (Guo et al., 2024).
Furthermore, a statistically significant relationship was confirmed between the location of the embryo within the cone and callus formation rate (χ2=11.68, df=2, P=0.0029). Immature embryos collected from the top and middle sections of the cones showed a higher formation rate than those collected from the bottom section. This suggests the presence of a physiological gradient within the cone. Embryos in the upper part of the cone may be in a more optimal developmental stage or have higher vitality, possibly containing more endogenous hormones (e.g., auxins and cytokinins) that are favorable for callus induction (Mulgund et al., 2012). This result clearly indicates that the position within the cone is an important factor to consider when establishing a tissue culture protocol for Korean fir. This finding is consistent with studies on other conifers, where the explant position and its corresponding physiological state are critical factors for culture success (Nawrot-Chorabik, 2008; Tautorus et al., 1991; Thorpe & Patel, 1986).
In conclusion, this study confirmed that several variables, including the collection site and location within the cone, collectively influence the success of callus induction in Korean fir. Our results provide a crucial foundation for developing a robust tissue culture protocol for this species, emphasizing the importance of selecting a healthy source material and optimizing the explant position. Further research should focus on comparing callus induction rates based on different medium compositions, which could lead to a better understanding of the underlying mechanisms governing callus induction.
Author Contributions
Conceptualization: HCP. Data curation: HCP. Formal analysis: IRK, DYP. Funding acquisition: HCP. Methodology: HCP. Project administration: HCP. Supervision: HCP. Validation: IRK, DYP. Visualization: IRK, DYP, DYL. Writing – original draft: IRK. Writing – review & editing: HCP, IRK.
Funding
This work was supported by a grant from the National Institute of Ecology (NIE) funded by the Ministry of Environment (MOE) of the Republic of Korea (NIE-B-2025-15).
Figures and Tables
Fig. 1
Map of Hallasan Mountain showing the four collection sites (Witse Oreum, Nambyeok, Nambyeok-Baengnokdam, and Baengnokdam) for Korean fir cones used in this study.
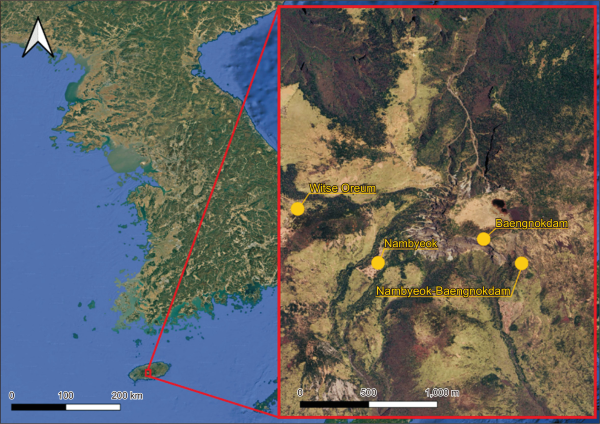
Fig. 2
Process of immature embryo isolation from Korean fir cones. (A) Appearance of Korean fir cones on the tree. (B) Freshly collected cones from Hallasan Mountain. (C) Cross-section of a cone showing the arrangement of seeds within. (D) A single seed with the outer and inner seed coats partially removed. (E) An isolated immature embryo after removal of the seed coats. (F) The final step of the process, showing the plated immature embryos on a medium.

Fig. 3
Sectional divisions of Korean fir cones from Hallasan Mountain. (A) The top section of the cone. (B) The middle section of the cone. (C) The bottom section of the cone.
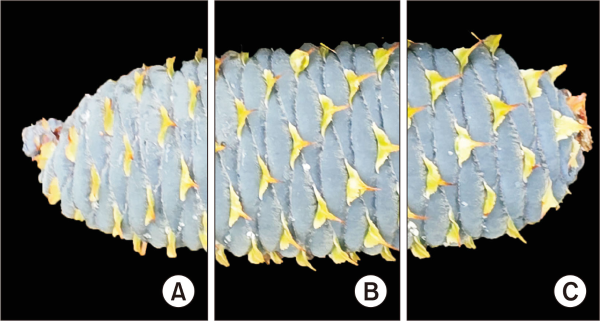
Fig. 4
Morphology of embryogenic callus derived from immature embryos. (A) Side view of the initial callus induced from the immature embryo. (B) Front view of the developing callus. (C) A later stage of the callus showing further development and formation of cell aggregates.
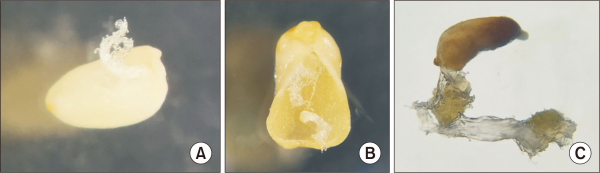
Fig. 5
Observed and expected counts of Korean fir callus formation from chi-square test. Bar plot of callus formation counts by cone position.
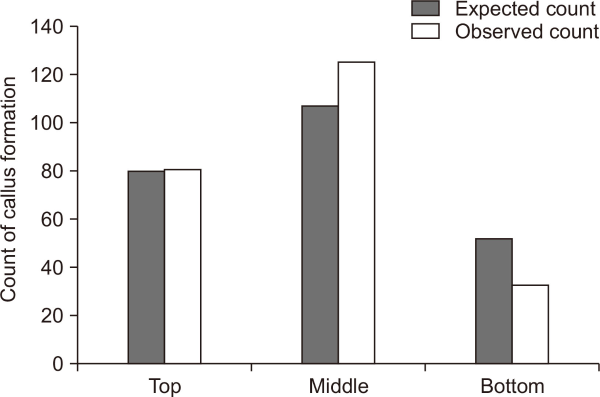
Table 1
Coordinates of Korean fir cone collection sites
| Collection site | Latitude (N) | Longitude (E) |
|---|---|---|
| Witse Oreum | 33°21’58.7” | 126°31’03.1” |
| Nambyeok | 33°20’52.8” | 126°31’11.2” |
| Nambyeok-Baengnokdam | 33°20’44.5” | 126°31’18.2” |
| Baengnokdam | 33°20’51.5” | 126°31’18.2” |
Table 2
Number of seeds plated by collection site, cone position, and media composition
| Collection site | Cone position | Medium | |||
|---|---|---|---|---|---|
|
|
|||||
| A | B | C | D | ||
| Witse Oreum | Top | 66 | 66 | 66 | 54 |
| Middle | 48 | 48 | 54 | 54 | |
| Bottom | 24 | 23 | 30 | 49 | |
| Nambyeok | Top | 16 | 12 | 12 | 12 |
| Middle | 18 | 12 | 20 | 24 | |
| Bottom | 0 | 6 | 6 | 2 | |
| Nambyeok-Baengnokdam | Top | 84 | 78 | 80 | 87 |
| Middle | 129 | 162 | 140 | 156 | |
| Bottom | 71 | 45 | 78 | 55 | |
| Baengnokdam | Top | 12 | 12 | 6 | 6 |
| Middle | 6 | 12 | 6 | 6 | |
| Bottom | 12 | 12 | 16 | 6 | |
Table 3
The number of samples with callus formation
| Collection site | Cone position | Medium | |||
|---|---|---|---|---|---|
|
|
|||||
| A | B | C | D | ||
| Witse Oreum | Top | 0 | 2 | 0 | 0 |
| Middle | 2 | 2 | 0 | 0 | |
| Bottom | 0 | 0 | 0 | 0 | |
| Nambyeok | Top | 0 | 0 | 0 | 0 |
| Middle | 0 | 0 | 0 | 0 | |
| Bottom | 0 | 0 | 0 | 0 | |
| Nambyeok-Baengnokdam | Top | 14 | 23 | 19 | 23 |
| Middle | 17 | 25 | 45 | 35 | |
| Bottom | 4 | 6 | 16 | 7 | |
| Baengnokdam | Top | 0 | 0 | 0 | 0 |
| Middle | 0 | 0 | 0 | 0 | |
| Bottom | 0 | 0 | 0 | 0 | |